t-cell receptor
Antigen-recognition molecules of the t-cell. Solely membrane bound, unlike b-cell receptor.
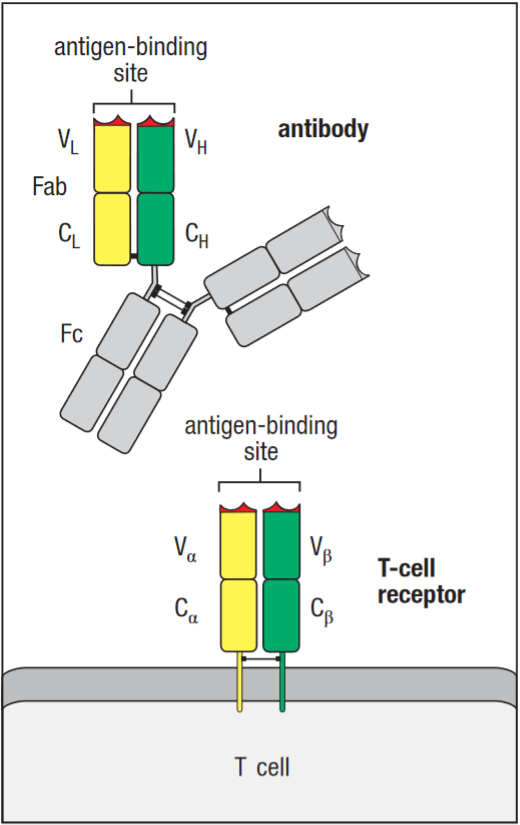
They are related to immunoglobulins in their structure with variable and constant regions, and the process of generation (see TCR V(D)J recombination).
However, unlike the b-cell receptor it does not recognise the antigen by itself, instead it recognises short peptide fragments and the MHC molecule it is bound to. This introduces an extra dimension to antigen recognition by T-cells known as MHC restriction - any given TCR is specific for a particular peptide bound to a particular MHC molecule.
The α and β chains are composed of two Ig domains each, and are responsible form antigen binding in most t-cells.
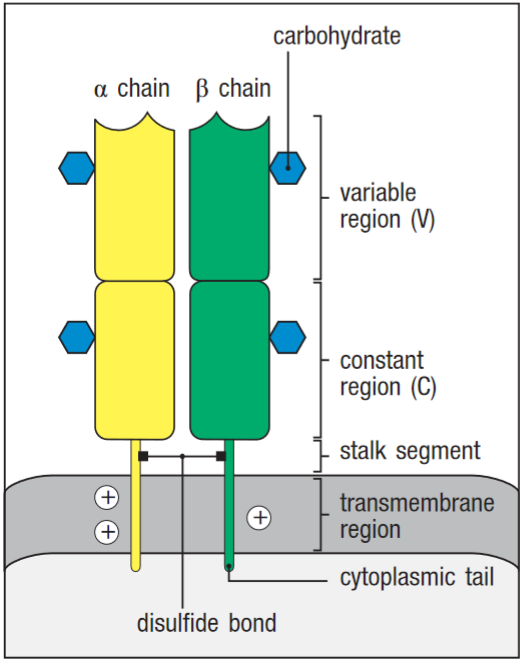
The TCR heterodimer is composed of two transmembrane glycoprotein chains. The transmembrane helices of both chains are unusual in containing positively charged residues.
TCR vs Fab domain in an Antibody
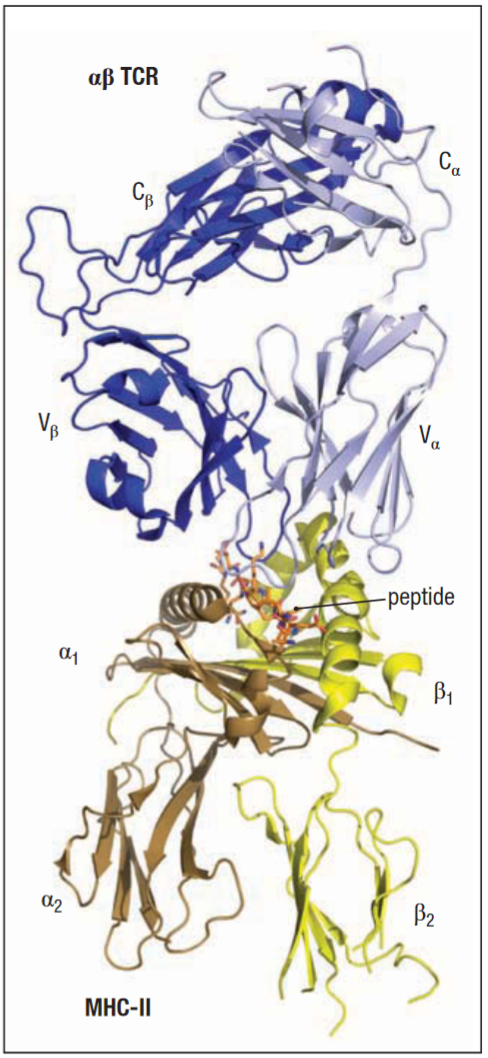
Interface of V and C domains in the TCR chains is more extensive than in Fab. Cα and β interaction may be assisted by carbohydrates with hydrogen bonding between the sugar in the glycosylation site and the Cβ domain. TCR CDR loops are also similar to antibodies, however there is some displacement when compared. Notably in the Vα CDR2 loop, which is oriented at roughly right angles to the equivalent loop in the antibody V domains, this also occurs in some Vβ CDR2 loop domains. These differences influence the ability of the TCR to recognise their specific ligands. Additionally, TCR has a fourth hypervariability region HV4, in both of its chains.
A TCR recognises antigen in the form of a complex of a foreign peptide bound to an MHC molecule.
γ:δ TCRs
A minority of t-cells bear a structurally similar TCR made up of γ and δ chains, and seem to have different antigen-recognition properties from the α:β TCRs.
Like α:β t-cells, γ:δ t-cells can be found in the lymphoid tissues of all vertebrates, but are also prominent as populations of intraepithelial lymphocytes, especially in the skin and female reproductive tracts, where their receptors display very limited diversity.
γ:δ t-cells do not generally conform by MHC restriction and can recognise their target antigens directly and thus are able to respond rapidly to molecules expressed by many different cell types.
Additionally, they can bind antigens presented by ‘nonclassical’ MHC class Ib molecules.
It has been suggested that they act as an intermediate between wholly innate immunity and adaptive immunity. Hence the role of transitional immunity clarifies the role of these t-cells.